We use cookies to make sure that our website works properly, as well as some optional cookies to personalize content and advertising, provide social media features and analyse how people use our site. Some countries outside of the European Economic Area do not have the same data protection standards as the country where you live, so if you accept some or all optional cookies, you give consent to the processing of your personal data, including transfer to third parties. You can find more information about how your personal data is processed when you click on theManage settings. Further information can be found in our privacy policy.
This article is about Nature. The soft tissues in the fossils have yielded important insights into the evolution of feathers. There is new evidence of feathers in pterosaurs, but it is not clear if they are related to the feathers in dinosaurs. Reports of pterosaur feathers with ovoid melanosome geometries suggest that they exhibited limited variation in colour. There are different melanosome geometries in the skin and feathers of a tapejarid pterosaur found in Brazil. In theropod dinosaurs, the melanosomes form distinct populations in different feather types and skin. The function of feathers in visual communication has deep evolutionary origins. The features show that early in feather evolution, genetic regulation of melanosome chemistry and shape 7 DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch DropCatch The success of birds 10 and the diversity of non-avian dinosaurs from the Middle Jurassic onwards are linked to feather innovations. The early evolutionary history of feathers is controversial as relevant fossils are rare. The pycnofibres in pterosaurs were recently dubbed feathers on the basis of preserved branching, but their function remains unclear. The small size and lack of secondary branching precludes functions in active flight, but their dense packing and distribution over the body are consistent with thermoregulation. Functional hypotheses for small, simple feathers in theropod dinosaurs are related to this. Simple unbranched feathers in theropods functioned in visual signalling, as evidenced by melanosome-based colour patterning. It's not clear whether feathers in earlier-diverging taxa also functioned in patterning. The timing and context of the evolution of melanin-based colour patterning in feathers is unknown. There is evidence of colour patterning, but this could be a taphonomic artefact. The feather colour in birds 16 is linked to variation in the melanosomes. There have been previous observations of feather melanosomes in pterosaurs. These are similar to melanosome geometries in the skin of extant reptiles and preserved melanosomes in the skin of fossil non-dinosaurian reptiles. The ability to vary melanosome geometry and control the colour of appendages is unique to theropods. Variable melanosome geometries in mammals suggest earlier origins for this feature in a common amniote ancestor and a secondary loss in pterosaurs. This issue is solved using a new specimen of an adult tapejarid pterosaur from the Lower Cretaceous Crato Formation 17 in Brazil. The incomplete cranium is associated with preserved skin, monofilaments and branched integumentary structures. Birds 18 and theropod dinosaurs were the only dinosaurs to have melanosomes that show tissue-specific geometries. The results show that feathers in pterosaurs are critical for tissue-specific partitioning of melanosome geometry and that it has deep evolutionary origins. There is a preserved soft tissue crest on the incomplete cranium. The white arrowhead indicates the calamus. 50mm scale bars, 5mm scale bars, 2mm scale bars, and 250mm scale bars. There is a new specimen of Tupandactylus that is preserved on five limestone slabs. The left nasoanteorbital fenestra, the occipital process, and the part of the cranium that is the anterior portion are all present. Between the postpremaxillary and occipital processes, there is a preserved soft tissue cranial crest. There are two types of integumentary structure close to the occipital process. The portion of the occipital process that is associated with monofilaments is 30mm long and 90mm wide. 1 and 2 These are feathers from stage I of the anurognathid Jeholopterus ningchengensis, Sordes pilosus and 24 juvenile anurognathids. The part of the occipital process associated with branched integumentary structures is called the distal part. There is a poorly defined central shaft that thins close to the tip. The portion of the shaft that resembles a calamus is narrow and light-toned. A branched structure is formed by the straight and closely spacing secondary fibres extending from the shaft. The branched structures can be straight but are often curved. Only when the central shaft and the branches are stiff and the branches are not diverging from a single point can the splaying be generated. This mode of branching is similar to the one in stage IIIa feathers 19 and 20 of the birds. There is strong evidence that the structures are feathers. This supports recent claims of branched feathers in other pterosaurs. The monofilaments are most likely to be stage I feathers. Stage IIIa feathers have never been reported in pterosaurs. The Tupandactylus branched structures are similar to the dromaeosaurid dinosaur Sinornithosaurus millenii 27 but different from the three types of branched feathers described in anurognathid pterosaurs 2. Branching in the anurognathid feathers can be either near the center or near the edge. The branched feathers in Tupandactylus branch are almost the entire length of the rachis, unlike the three anurognathid feather types. The length of the Tupandactylus secondary fibres differ from the length of the anurognathid feathers. The Tupandactylus feathers are not taphonomic. There are multiple feather types in the specimen, which is consistent with the presence of anurognathids 2, feathered dinosaurs 29 and 30 and fossil 32 and 33 and extant birds 34. Tupandactylus includes many isolated feathers where branching is obvious and thus cannot be explained by superposition. The consistency of branching feathers is not reflected by the random pattern of degradation expected from decay. The feathers are restricted to a portion of the skull that should be devoid of structural fibres of the skin that have decayed. The cranial crest lacks feathers despite the preservation of long straight fibres that are up to 300mm long. 1, 4 A recently published phylogeny for pterosaurs, birds and non-avialan dinosaurs preserved integumentary structures. Tupandactylus has no secondary branching, so their branched feathers correspond to an open pennaceous vane. Ancestral-state estimates show that the most likely result is that the avemetatarsalian ancestors of pterosaurs and dinosaurs possessed integumentary filaments. The Extended Data Table 2 has a number of items. There is a hypothesis that the integumentary structures of both groups originated independently. The more parsimonious interpretation is that feather morphologies have a single origin. The model predicts that there were more complex integumentary structures within Theropoda. The Extended Data Table 2 has a number of items. This doesn't mean that feather types in each group are the same. Some feather morphologies are shared, but others are not, for example, feathers with barbules in theropods and pterosaurs. Barbules are an innovation of theropod feathers. The younger age of Tupandactylus is similar to the older age of anurognathids, with branching restricted to a narrow zone on the shaft. The results of ancestral-state estimations for the origin of feathers with the highest likelihood are shown in the phylogeny. The most complex structure is shown for each taxon. The ancestors of dinosaurs and pterosaurs had feathers. The best model that estimates trait transition rates following ordered evolution is the mbl branch length estimation. The scaled likelihoods of different integumentary structures are shown in the pie charts. The values for model parameters are shown in the table. The silhouette of Tupandactylus is drawn by E. Boucher. The Silhouettes of integumentary appendages are reproduced. Springer Nature limited. We analysed samples of soft tissue from the cranial crest. The data table shows that all soft tissue samples contain abundant ovoid or elongate microbodies. The degraded remains of the feather matrix can be seen in the microbodies embedded in the matrix. Microbodies are restricted to the soft tissues in samples 1 and 9 that are adjacent to the cranial crest. The crest of a Tupandactylus specimen from the Crato Formation 5 and a pterosaur from the Jehol Group 6 have previously been reported with relatively homogeneous ovoid geometries. The preserved melanosomes 5, 6 were interpreted as the microbodies. This is consistent with the broad consensus that similar microbodies, preserved in dark carbonaceous soft tissue films associated with other fossil vertebrates, are fossils. In Tupandactylus, melanosomes from the skin fibres in the crest, monofilaments and branched feathers differ significantly in geometry. The melanosomes are restricted to the monofilaments. The ovoid in the branched feathers are long and wide. There are ovoid skin fibres located between the base of the cranial crest and the occipital process. The melanosomes are spheroidal in the crest. Similar tissue-specific partitioning of melanosome geometry has been reported in other fossils. The other specimen 5 of Tupandactylus has no melanosome populations. The reported diversity of melanosome morphologies expands the known range 2, 6 of geometries of pterosaur melanosomes. The geometry of the melanosomes in Tupandactylus is similar to that of other animals. This supports the idea that pterosaurs are made of feathers. It does not completely exclude the alternative hypothesis that pterosaurs have a third type of integumentary outgrowth that is capable of giving different colors. The melanosomes in the feathers are suggestive of different colors. The data confirm tissue-specific melanosome populations regardless of the colour produced. This strongly suggests that the genes needed for tuning melanosome geometry were already present in the avemetatarsalian ancestors of pterosaurs. Our study has implications for understanding the evolution of colouration. Melanosomes in other pterosaur fossils have ovoid to spheroidal shapes. This low melanosome diversity is similar to the skin of lizards, where many colors are generated by non-melanin pigments. The preservation of ovoid and spheroidal melanosomes in the feathers and skin of pterosaurs was seen as evidence for the retention of the ancestral state. There are important differences in melanosome geometry between the skin and feathers, with evidence for expanded diversity of melanosome geometry in the feathers. This tissue-specific partitioning of melanosome geometry characterizes birds and mammals. This feature may reflect preferential selection of more extreme, oblate melanosome geometries in order to expand colour space 40 into regions associated with eumelanin-dominated darker and iridescent hues. This may be a response to the loss of chromatophores during the evolution of integumentary appendages. Changes in metabolism 6 and immunity 40 may be the cause of the fundamental changes in skin structure. The production of eumelanin-rich melanosomes reflects earlier activity of melanocyte-stimulating hormone 7 and/or enhanced production of premelanosome proteins. The discovery of melanosome in the feathers, but not the skin, of Tupandactylus expands the known range of melanosome geometries in pterosaurs. The timing of the origin of the regulatory networks governing melanogenesis is linked to three evolutionary scenarios. The genotypic and phenotypic characters could either be ancestral to avemetatarsalians or evolved independently in theropods and pterosaurs. The most parsimonious scenario is that feathers in the avemetatarsalian ancestors had melanosomes. This feature is consistent with a single deep evolutionary origin, where critical shifts in the genetic machinery facilitating plasticity in melanosome shape occurred in the common ancestor of pterosaurs and birds. In the Middle to Late Triassic, key controls on the colouration of theropods and fossil and extant birds were already in place. Twenty-two soft tissue samples were collected using sterile tools. There are six distinct integumentary appendages located close to the occipital process. There are two samples of the matrix in the region between the cranial crest and the skull. Soft tissue samples were mounted on carbon tape and coated with gold. The working distance of the two microscopes was 8.6 and 13mm, respectively. A total of 2,994 melanosomes were measured using the long and short axis. For some samples, orientation was measured. The mean, standard deviation, skew and coefficient of variance were calculated for melanosome length, width and aspect ratio. The ANOVA test was used to determine the significance of variation in the data. The data on melanosome geometry was analysed using the MASS package 50 and the Nnet-package, both implemented in R. Ancestral-state estimations for integumentary appendages were done using the methodology and data. 2 The appendages were assigned to one of six categories: scales, monofilaments, brush-like filaments, tufts of filaments joined, open pennaceous vane lacking secondary branching and closed pennaceous feathers. The data on feathers from the MCT.R.1884 was included in the database 2. The function of the ape 4 package 52 was used for maximum-likelihood estimations. The tree branch lengths were estimated using two methods. See ref for more details. 2 Four evolutionary models with different state transition rates were used for our analyses. If open-vaned feathers have already evolved, feathers with a closed vane can only transition from one state to another. The model that corrects for sample size is summarized as weighed AICc values in theExtended Data Table 2. The large parameter space made it difficult to estimate ancestral states for the mbl-ARD model. The function of the phytools package 54 was used. The Nature Research Reporting Summary contains further information on research design. The data on melanosome geometry and the character matrix used in the analyses can be found in the Zenodo.org data repository. Images and samples from the corresponding authors are available on request. In The Evolution of Feathers, X. The Pterosaur integumentary structures have feather-like branching. Nat. Ecol. Evol. 3, 24 and 30 of 2019. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the No feathers on pterosaurs. Nat. Ecol. Evol. The year is 1590. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the The origin of feathers. The trends ecol. Evol. 34, 856 The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the F. L. Pinheiro and his colleagues studied the chemical characterization of pterosaur melanin. There is a science. Rep. 9, 15947 The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the The evolution of melatonin shows a key shift within feathered dinosaurs. Nature 509, 350,353. The article is from the Google Scholar. Genetics of hair and skin. Annu. The Rev. Genet. 37, 67, 90, 2003 The article is from the Google Scholar. Evolutionary shifts in the color system of birds. Evolution 70 was published in 2016 The article is from the Google Scholar. Marks, M. S., Raposo, G., and Melanosomes, dark organelles. Nat. Rev. Cell. 8, 797 in 2007. The article is from the Google Scholar. The origin and evolution of birds. It's a word. There is a Biol. 25, R888. The article is from the Google Scholar. There are no feathers on pterosaurs. Nat. Ecol. The evol. 4 was published in 2020. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the In Animal Adaptation to Cold was written by R. L. and W. R. The extinct dinosaur had Plumage color patterns. Science 327, 1373,1372 The article is from the Google Scholar. Countershading and stripes in the theropod dinosaur Sinosauropteryx reveal heterogeneous habitats. It's a word. There is a Biol. 27, 3343 The article is from the Google Scholar. The colour of fossil feathers. There is a Biol. Lett. There were 4, ,,,,,,,,,,,,,,,,,,,,,,,,,, The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the In Bird Coloration: Mechanisms and Measurements was written by K. J. The press, 2006). The Crato Fossil Beds of Brazil: Window into an Ancient World was written by D. M., G. and R. F. Press, 2007. The internal structure of extinct animals is revealed by the geometry and chemistry of modern melanosomes. Proc. The National Acad. There is a science. USA 116, 17880. The article is from the Google Scholar. The origin and development of feathers. J. Exp. Zool. There was a report in 1999 about a report called,,,,,,,,,,,,,,,,,,,,,,,, The article is from the Google Scholar. The evolution of feathers. Q. Rev. There were 77, 261, and 295 in 2002. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the The soft tissue of Jeholopterus and the structure of the pterosaur wing. Proc. R. Soc. Lond. B 277, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, 322, There is a scholar on the internet. A nearly completely articulated rhamphorhynchoid pterosaur with extremely well-preserved wing membranes and hair was found in Inner Mongolia. Chin. There is a science. Bull. There were 47, 226, and 230 in 2002. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the Sharov, A. G. phylogeny of the orthopteroidea was published in 1971. The nature of the pterosaur flight apparatus is discussed. Nature 372, 64 (1994). The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the An Early Cretaceous Heterodontosaurid dinosaur with integumentary structures. Nature 458, 333,336, 2009. The article is from the Google Scholar. A therizinosauroid dinosaur with integumentary structures was found in China. Nature 399, 350, and 355 were published in 1999. The article is from the Google Scholar. The Yixian Formation of China contains a dromaeosaurid dinosaur. b. The article is from the Google Scholar. Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature 410 was published in 2001. The article is from the Google Scholar. Four-winged dinosaurs from China. Nature 423, 335,340. The article is from the Google Scholar. A pre-Archaeopteryx troodontid theropod from China has long feathers on the metatarsus. Nature 465, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, 663, The article is from the Google Scholar. Godefroit, P. et al. describe a dinosaur from Siberia with both feathers and scales. Science 347, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, 431, The article is from the Google Scholar. Chiappe, Ji, S. A., Ji, Q. and Norell, M. A. studied the systematics of theropoda from northeastern China. Bull. Am. Mus. Nat. Hist. In 1999 The origin of feathers and a primitive bird. Science 290 was published in 1955. The article is from the Google Scholar. An overview of the integumentary of modern birds is provided by P. R. Stettenheim. Am. Zool. 40, 465, 475 (2000). There is a scholar on the internet. No feathers on pterosaurs. Nat. Ecol. Evol. The year is 1590. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the In The Evolution of Feathers: From their Origin to the Present is a book written by Campione, Barrett, P. M., and Evans. Fossilized melanosomes and the colour of dinosaurs and birds. Nature 465, 1075, 1078. The article is from the Google Scholar. The evolution of iridescent plumage and the reconstruction of Microraptor. Science 335, 1215, 1219, 2012 The article is from the Google Scholar. Alba, L. and Shawkey, M. D. Melanosomes: biogenesis, properties, and evolution of an ancient organelle. There is a science to it. Rev. 99 was published in 2019. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the M.E. McNamara and her co-authors decode the evolution of melanin in the body. The trends ecol. Evol. The year is 36, and it will end in 2021. The article is from the Google Scholar. Rogers, C. S. and colleagues studied the X-ray absorption of melanosomes in animals. Proc. R. Soc. B 286 was published in 2019. The article is from the Google Scholar. Hierarchical biota-level and taxonomic controls on the chemistry of fossil melanosomes revealed using synchrotron X-ray fluorescence. There is a science. Rep. 10, 8970 was re-elected in 2020. The article is from the Google Scholar. In Biology of the Integument was written by Landmann. Alexander, N.J. and Fahrenbach, W. H., studied the chromatophores of Anolis carolinensis. Am. J. Anat. There were 126, 41, and 55 in 1969. The article is from the Google Scholar. In Biology of the Reptilia was written by Cooper and Greenberg. The Chicago Press was published in 1992. A guide to the field of palaeo colour: reconstructing colour patterns from ancient organisms can give new insights to ecology. Bioessays 37 was published in 2015. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the The expression of Pmel17 reveals cranial melanoblast migration. Gene Expr. Patterns 3, 703, and 2003 The article is from the Google Scholar. M. D., Magalh, P. J., and Ram are all related. Int. of biophotonics. 11, 36, 42, and 11 were all published in 2004. There is a scholar on the internet. The PAST: paleontological statistics software package is used for education and data analysis. There is a Palaeontologia Electr. There were 4, 9 and 4 in 2001. There is a scholar on the internet. Venables, W. N. and Ripley, B. D. Modern Applied Statistics with S. Characterization of melanosomes involved in the production of non-iridescent structural feather colors and their detection in the fossil record. J. R. The interface 16 was published in 2019. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the There is an analysis of phylogenetics and evolution with R. Bell, M. A. and Lloyd, G. T. Strap are the authors of an R package for plotting phylogenies. Palaeontology 58, 389. The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the L. J. Phytools is an R package for comparative biology. There are methods Ecol. Evol. 3, 217,223, 2012 The article was written by a scholar from the internet search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the search engine, the Download references The work was funded by a grant from the Irish Research Council. T. Hubin was the photographer of MCT.R.1884. The authors do not have competing interests. Nature thanks Michael, Stephen, and Xing for their contribution to the peer review of this work. There are peer reviewer reports available. The soft tissue crest is brown. The dark part of the crest is darker than the rest of the crest and the brown fibres are not visible in that area. The scale bar is 100mm. A schematic illustration of a project. The region immediately adjacent to the occipital process and the branched feathers are restricted. The preserved bones are shown in white while the cranial soft tissue crest is shown in dark grey. The black part of the skull is not present on the slab. The image was taken by Julio Lacerda. The scale bar is 100mm. Only scenario 3, with a stiff central shaft and stiff barbs of equal length, can explain the particular structures observed in Tupandactylus feathers. The scale bars are 1mm. The Ventral part of the soft tissue crest is separated from the occipital process by a zone lacking soft tissue. The brown fibres of the soft tissue crest are oriented in a different way than the rest of the specimen. The scale bars are 10 mm (a, d); 2 mm (b, c); and 5 mm (e, f). The values for model parameters are shown in the table. There are different types of integumentary structures, including scales, monofilaments, brush-like filaments, tufts of filaments joined basally, open pennaceous vane lacking secondary branching and closed pennaceous feathers. The silhouette of Tupandactylus was created by Evan Boucher. The Silhouettes of integumentary appendages are reproduced. 2 There is a fig. The values for model parameters are shown in the table. There are different types of integumentary structures, including scales, monofilaments, brush-like filaments, tufts of filaments joined basally, open pennaceous vane lacking secondary branching and closed pennaceous feathers. The silhouette of Tupandactylus was created by Evan Boucher. The Silhouettes of integumentary appendages are reproduced. 2 There is a fig. The values for model parameters are shown in the table. There are different types of integumentary structures, including scales, monofilaments, brush-like filaments, tufts of filaments joined basally, open pennaceous vane lacking secondary branching and closed pennaceous feathers. The silhouette of Tupandactylus was created by Evan Boucher. The Silhouettes of integumentary appendages are reproduced. 2 There is a fig. The branched feathers contain melanosomes from the soft tissue crest. The scale bars are 2 BC. Data from refs, Melanosome geometry in amniotes. This study is about mammal hair 6 and the skin of Pterosaurs. There is no data from non-avialan dinosaurs here. The range of melanosome geometries known for bird feathers can be seen in the picture. Darker shades indicate more than one data point with the same measurement. There are three different categories of melanosome geometry. Only taxa for which melanosome length and aspect ratio was known have been included. The silhouette of Tupandactylus was created by Evan Boucher. There are references in this file. You agree to abide by the Community Guidelines when you submit a comment. If you find something that does not comply with our guidelines, please flag it as inappropriate.

Scanning electron microscopy
Measurements of melanosome geometry
Ancestral-state estimations
Reporting summary
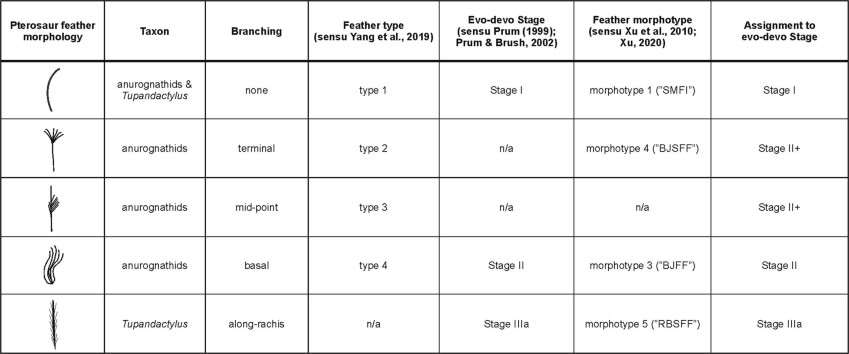
There is an assignment of pterosaur feathers, including those reported in Tupandactylus. There are three types of integument: SMFI: slender monofilamentous integument, BJFF: slender monofilamentous integument, and BJSFF: slender monofilamentous integument.
All Rights Reserved © 2024